Пути гематоинтерстициального транспорта
Поскольку физиологическая теория пор основана на представлении о простой геометрической форме транспортных каналов в эндотелии — цилиндрической или щелевидной — у морфологов появилась надежда визуализировать эти гипотетические пути с помощью электронного микроскопа и метода «молекулярных зондов».
Для установления соответствия между величиной тест-молекул (а, следовательно, и размеров пор) и пропускной способностью реальных «каналов» в эндотелии микрососудов были широко использованы трассеры биологического происхождения: гемсодержащие пептиды, известные под коммерческим названием микропероксидазы (мол. масса 1500—1900 дальтон, диаметр молекулы 2 нм), цитохром с (мол. масса 12000, диаметр молекулы 3,3 нм), миоглобин (мол. масса 17 800 дальтон, размеры молекулы 2,5X3,4X4,2 нм), растительная пероксидаза (мол. масса 40Х103 дальтон, диаметр молекулы 5 нм), гликоген (диаметр молекулы 20—30 нм), декстраны (мол. масса 7500—25 000 дальтон, диаметр молекулы 12,5—22,5 нм), гемоглобин (мол. масса 68Х103 дальтон, размеры молекулы 6,4X5,5X5,0 нм), ферритин (мол. масса 450Х103 дальтон, диаметр молекулы 11 нм) [Буланова Г. В., Сапин М. Р., 1977; Караганов Я. Л. и др., 1980, 1981; Karnovsky М., 1967; Bruns R., Palade G., 1968; Simionescu N, Palade G., 1971; Simionescu N. et al., 1972, 1973, 1978; Wissig S., Williams M., 1978; Wissig S, 1979; Welt K. et al, 1980, 1981].
На основании результатов электронно-микроскопических исследований предложено выделять три группы путей трансэндотелиального транспорта [Чернух А. М, 1979; Караганов Я. Л. и др., 1978]: парацеллюлярные (межклеточные), трансцеллюлярные и комбинированные, т. е. сочетающие пути транспорта жидкости между клетками и через них.
Парацеллюлярными путями служат контакты между клетками. Последние представляют собой специализированные образования, которые возникают благодаря взаимодействиям краев смежных эндотелиальных клеток или их отростков. На ультратонких срезах межклеточные каналы выглядят в виде щелевидных пространств между цитолеммами соседних эндотелиоцитов, посредством которых просвет обменных микрососудов сообщается с субъэндотелиальным пространством. Результаты количественных исследований показали, что на долю межклеточных щелей приходится от 0,08 до 0,2% внутренней поверхности эндотелия капилляров [Simionescu М. et al, 1974].
«Микролимфология», В.В.Купирянов, Ю.И. Бородин

Так, уже через 2 мин после внутривенного введения водного раствора ферритина его молекулы заполняли кавеолы, открывающиеся на люминальной поверхности эндотелиоцитов, а через 10 мин ферритин обнаруживался в соединительной ткани, окружающей капилляры [Bruns R., Palade G., 1968]. N. Simionescu и соавт. (1973) использовали в качестве трассера миоглобин и зарегистрировали, что в первые 30—35 с после его…

Эта точка зрения была основана на изучении препаратов, обработанных в процессе подготовки для электронно-микроскопического изучения таниновой кислотой. Известно, что последняя обладает сродством к мембранным белкам, а также эпизодически интенсивно метит покрытия клеток. Отложения таниновой кислоты встречаются всюду, где имеется свободный доступ к поверхности клеток [Wagner R., Casley-Smith J., 1981]. Использование этого методического подхода позволило продемонстрировать не…

Считается, что цитолемма эндотелиальных клеток малопроницаема для молекул, растворимых в воде [Folkow В, Neil Е, 1976]. Поэтому утечка последних должна происходить преимущественно по парацеллюлярным шунтам — межклеточным каналам в эндотелиальном слое обменных микрососудов (смотрите рисунок ниже). Открытый канал (указан стрелкой) (Э1 и Э2) в стенке кровеносного капилляра между эндотелиальными клетками Почка крысы.…

Характерная черта коммуникационных контактов (синоним: gap junctions) — кластерное распределение внутримембранных частиц (смотрите рисунок ниже). Коммуникационные контакты эндотелиальных клеток венулы Разлом клеточной мембраны обнажил внутримембранные частицы, расположенные в виде кластеров (указаны стрелками) на Р-поверхности цитолеммы. Миокард мыши. Криофрактограмма. X 60 000 (препарат Я. Л. Караганова и Г. А. Алимова). На Р-поверхности цитолеммы такие агрегаты представлены…

На основании данных, полученных методом криоскалывания, считается, что в замыкающих контактах внутримембранные глобулярные частицы располагаются линейно, образуя сеть «гребешков» на Р-поверхности и комплементарных им «желобков» на Е-поверхности цитолеммы (смотрите рисунок ниже). Замкнутая сеть контактных гребешков (указаны стрелками) на Р-поверхности скола цитолеммы эндотелиальных клеток кровеносного капилляра Миокард мыши. Криофрактограмма. X 35 000 (препарат Я. Л. Караганова…
Если одни исследователи [Karnovsky М. 1967; Wissig S, Williams М.. 1978; Wissig S, 1979] склоняются к положительному решению этого вопроса, то другие [Simionescu N. et al, 1975; Palade G. et al, 1979] в качестве эквивалента мелких пор рассматривают так называемые трансэндотелиальные каналы, образованные цепочками слившихся между собой микропиноцитозных везикул. Одна из причин неоднозначного соответствия мелких…

В качестве трансцеллюлярных путей описаны плазмалеммальные везикулы и фенестры. Плазмалеммальные (микропиноцитозные) везикулы Плазмалеммальные (микропиноцитозные) везикулы являются наиболее характерными элементами цитоплазмы эндотелиальных клеток и на ультратонких срезах представляют собой сферические и эллипсоидные образования, ограниченные элементарной мембраной [Шахламов В. А, 1971; Караганов Я. Л., 1972; Чернух А. М. и др., 1975; Palade G, 1953]. Везикулы бывают двух типов:…

Стереологические особенности, топография и лимитирующие размеры везикулярных каналов позволили идентифицировать их в качестве структурного эквивалента мелких пор [Simionescu N. et al., 1975; Palade G. et al., 1979]. Позже было показано, что через трансэндотелнальные каналы проникают не только микропероксидаза, но и растительная пероксидаза [Караганов Я. Л. и др., 1980, 1981; Welt К. et al., 1980, 1981],…
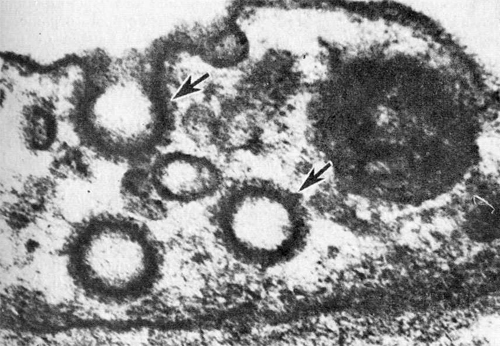
Благодаря использованию катионизированного ферритина удалось показать, что трансэндотелнальные каналы и диафрагмы, перекрывающие устья плазмалеммальных везикул, практически лишены анионных участков [Simionescu N., 1979]. Дальнейшая разработка вопроса об электрохимических различиях трансцеллюлярных путей эндотелия, проведенная с помощью перфузии сосудистого русла растворами лектинов, связанных с растительной пероксидазой или с ферритином, и топографическим анализом Мест связывания меченых лектинов позволила продемонстрировать, что…
Функциональное значение окаймленных везикул в настоящее время связывают со специфическим поглощением макромолекул посредством процесса, получившего название «рецепторопосредованного эндоцитоза» [Anderson R. et al., 1976; Goldstein J. et al., 1979]. При этом локализацию клеточных рецепторов1 связывают с зонами окаймленных углублений (coated pits), которые после отшнуровки от цитолеммы превращаются в рофеосомы. Если учесть сведения о существовании на поверхности…